
HLA基因分型介绍
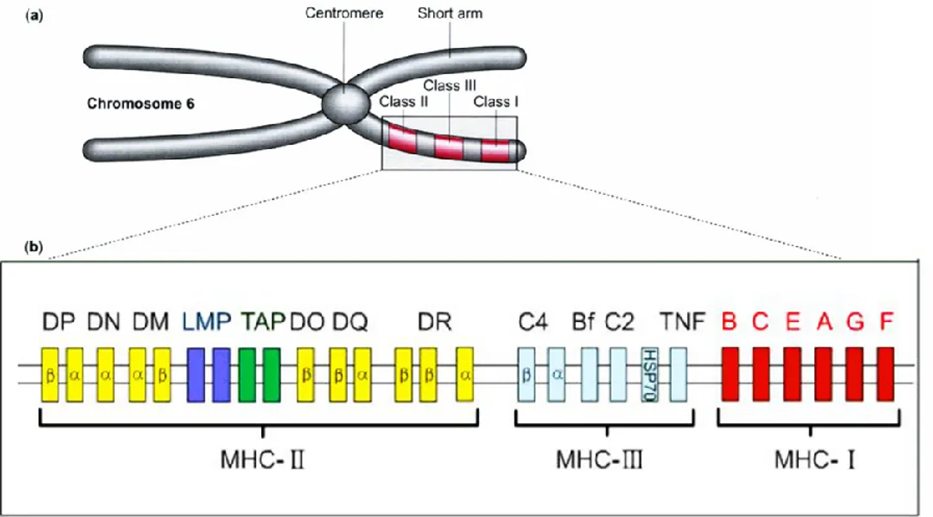
人类白细胞抗原(human leukocyte antigen)是在6号染色体短臂上的一类基因能够编码MHC蛋白。HLA是一种识别人类白细胞的重要的分子标志,能够使免疫系统区分本身和异体物质,具有非常重要的生物学功能。每个人的HLA千差万别,很少能有两个人存在完全相同的基因型,HLA有不同的基因座,编码两大类MHC蛋白。HLA在医学上的意义主要体现在器官移植,输血,疾病相关性,生理学等方面。
HLA基因结构
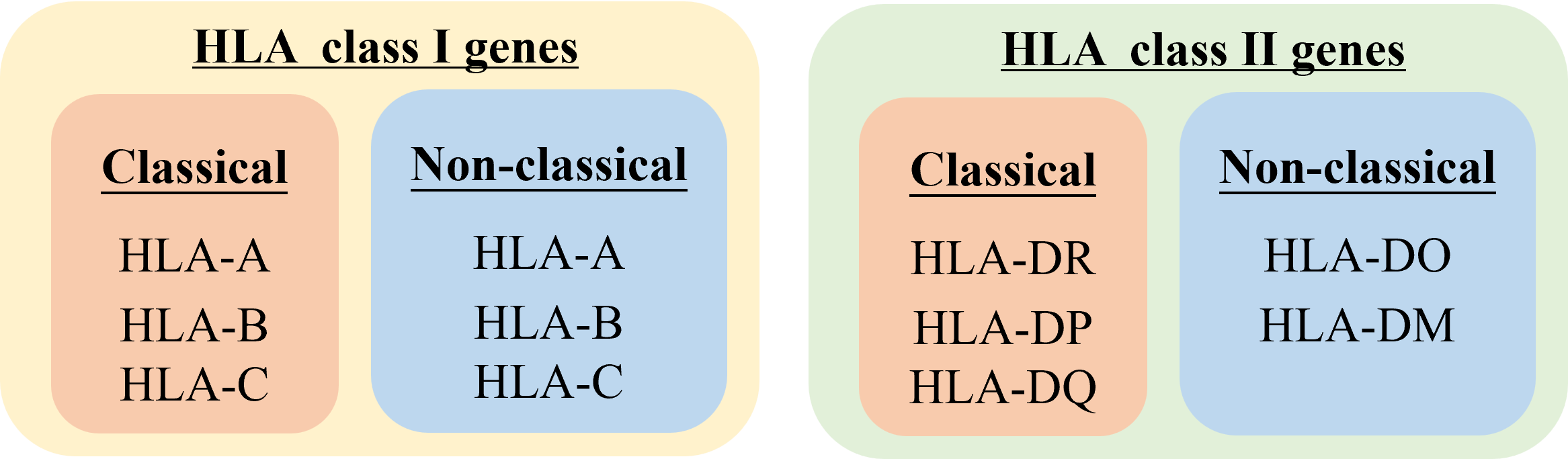
HLA复合体有224个基因座(locus),按其产物的结构、分布与功能分为两群。
HLA-I
经典I类基因: HLA-A、-B、-C参与递呈内源性抗原。
非经典I类基因: HLA-E、-G、-F。
HLA-I编码第一型MHC,能够将细胞内的抗原呈递到细胞表面,CD8+ T细胞(killer T cell)识别被感染细胞并消灭,与排异反应的发生密切相关。HLA-Ⅰ类抗原由重链(α链)和轻链(β链)经非共价键连成异二聚体。 α链由 HLA 基因编码, β链则定位于 15 号染色体。α链由三个胞外区(α1、α2和α3)、穿膜区和胞质区组成。α1 和α2组成抗原结合部位,α3 则是与 T 细胞表面的 CD8 分子结合的部位。HLA-I类抗原分布相当广泛,见于所有有核细胞,在淋巴细胞上密度最高,心肌或肝细胞在正常情况下极少或没有。血浆中有可溶性 HLA - I 类抗原,可能是细胞膜新陈代谢所致。成熟的红细胞上无 HLA 抗原,血小板上带有 HLA - I 类抗原。I类主要组织相容性复合体(MHC class I)抗原的处理和呈递过程:1. 肿瘤特异性突变产生的突变蛋白会被蛋白酶体降解为8~11aa的肽段;2. 这些肽段被抗原加工相关转运体(TAP)转运进入内质网腔;3. 与新合成的MHC-I结合;4. 最终通过高尔基体转运至细胞膜被CD8+ T细胞识别。HLA-Ⅰ类抗原表达异常在小鼠及许多人类肿瘤或肿瘤衍生的细胞株均已发现MHC-Ⅰ类抗原表达缺失或密度降低。若将Ⅰ类基因转染给肿瘤细胞株,则恶变细胞可发生逆转,且浸润性与转移性消失或降低。这可能是由于MHC-Ⅰ类抗原缺失的肿瘤细胞不能补TC识别并攻击,从而导致肿瘤免疫逃逸(sneaking through)。
HLA-II
经典II类基因: HLA-DP 、-DQ、-DR参与递呈外源性抗原。
非经典II类基因: LMP、TAP、HLA-DM、HLA-DO 参与抗原的加工和转运。
HLA -II型(HLA-DP, HLA-DM, HLA-DO, HLA-DQ, HLA-DR)编码第二型MHC,HLA- Ⅱ类抗原同样由α链和β链以非共价键组成的异二聚体。不同的是:两条链均由 HLA 基因编码,各有两个胞外结构区(α 1 、α 2 和β 1 、β 2 )、穿膜区和胞质区。α 1 与β 1 组成抗原结合槽,α 2 和β 2 则与 T 细胞表面的 CD4 受体结合。将细胞外抗原呈递给CD 4+T(T-helper cells 辅助T细胞),T细胞增殖,进而刺激B细胞产生抗原特异性抗体(体液免疫)。像是组织中有细菌侵入,则巨噬细胞进行吞食后,把细菌蛋白加工成肽链,通过MHC展示给辅助性T细胞,启动体液免疫反应。因为涉及细胞外抗原,所以只有吞噬性的细胞(比如巨噬细胞、B细胞)的表面有第二型MHC。HLA-ll基因主要与免疫反应的发生相关。自身抗原会被调节T细胞抑制(regulatory T cell)。HLA -Ⅱ类抗原分布较窄,密度最高的是树状突细胞、单核细胞、一些吞噬细胞亚群及 B 淋巴细胞。HLA分子对T细胞在胸腺内的分化成熟过程也起重要作用。体外研究发现:去除胸腺中MHCⅡ类抗原阳性的基质细胞,则CD4+ T细胞的发育受阻,在胸腺培养细胞中加入抗MHCⅡ类抗原的单克隆抗体,也能阻止CD4+ T细胞的发育。目前认为MHC分子在T细胞自身耐受的形成和T细胞库的产生中都起着重要作用。
HLA- Ⅱ类分子表达异常与自身免疫病的发生有关。器官特异性自身免疫疾病的靶细胞可异常表达HLA-Ⅱ类抗原。诸如Graves病患者的甲状腺上皮细胞、原发性胆管肝硬化患者的胆管上皮细胞、Ⅰ型糖尿病患者的胰岛β细胞等均可发现HLA-Ⅱ抗原异常表达。其机制可能是局部感染诱生IFN-γ,后者诱导Ⅱ类抗原表达。Ⅱ类抗原乃抗原呈递的效应分子,一旦靶细胞异常表达Ⅱ类抗原,就可能以组织特异性方式把自身抗原呈递给自身反应性T细胞,从而启动自身免疫反应。激活的自身反应性TH又可分泌大量IFN-γ,诱导更多的靶细胞表达Ⅱ类抗原,加重和延续自身免疫反应,最终导致迁延不愈的自身组织损伤。
HLA-Ⅲ
包括编码补体C4、Bf、C2的基因。
编码炎症相关分子TNF、HSP70等基因。
会产生某些补体、细胞因子以及热休克蛋白等,但不是主要的功能
HLA-I和II不同基因座位上分别编码的基因产物,称之为HLA复合体抗原,表达在细胞表面,呈凸起状。HLA的生物学功能不仅限与诱导同种异体组织或器官移植排斥反应相关,而且参与免疫应答的调控,如MHC限制性、抗原转递、补体产生等。HLA研究中,模拟免疫反应激活的过程,就要经过蛋白酶体对抗原(蛋白)的剪切预测、肽段转运、肽段和MHC结合亲和力预测以及T细胞识别预测等几个重要步骤。一般可以采用滑动窗口(sliding windows)的方法把可能的肽段都提取出来,MHC结合亲和力预测由软件实现(但效果各有说法,最好多重比较筛选,软件对比),更严谨的可以用抗原实验来验证(制造目标肽段,做实验查看与MHC的结合情况)。
HLA 配型
HLA配型也叫移植配型,HLA基因座位点具有高度多态性,完全相合配型的难度非常高。在HLA配型方面,主要进行HLA-A、HLA-B和HLA--DR三对位点的配型,只有两个个体的HLA配型完全相同才能进行造血干细胞移植,否则可能会发生两种情况,一是病人体内的免疫细胞把植入的供体细胞当作"异物"或"入侵者"进行攻击,称为"移植排斥反应",其结果是移植失败,"造血"不能在病人体内植活。另一种可能是供体的造血细胞在病人体内植活,产生大量的免疫活性细胞,这些细胞"反客为主"把病人的组织和细胞当作"异物"和"入侵者"进行攻击,最容易受攻击的组织和器官是皮肤、肝脏和肠道,发生皮疹、黄疸、转氨酶升高和腹泻不止甚至血便,称为移植物抗宿主病(GVHD),严重者可致命。一般血缘关系近的人,HLA配型相似度更高。”理论上说,同胞兄弟姐妹中有1/4的机会HLA配型完全相合;子女与父母之间只有一半HLA抗原相同,医学上叫做半相合。通常不能相互移植。所以骨髓移植最初主要在同胞兄弟姐妹之间进行。非血缘关系的hla位点相合率非常低MHC。
HLA 抗原和等位基因命名
座位名*4个数字 (前两位是指这个等位基因相应的血清学特异性;后两位数则代表该等位基因序号。如果两个等位基因的4个数字不同,则这两个等位基因编码的蛋白不同。),HLA配型需要细分类完全相同。
用血清学及细胞学技术检测的抗原特异性 HLA抗原的检出最初采用诺贝尔奖获得者法国人Dausset倡导的白细胞凝集反应,随后由荷兰人VaT/Rood、美国人Terasaki等建立了血清学分型技术,即补体依赖的微量淋巴细胞毒试验。其通用的标准方法称为NIH二步法。HLA-A抗原、HLA-B抗原、HLA-C抗原及HLA-DR抗原、HLA-DQ抗原可用血清学方法分别检出,其检出的抗原特异性覆盖面较广,称为宽特异性。
用分子生物学方法检测的等位基因 20世纪80年代后期,分子生物学引入HLA领域,并进一步在PCR基础上发展了各种DNA分型技术,称为基于PCR的HLA基因分型(PCR-based HLAgenotyping)。常用有PCR-RFLP(PCR扩增产物的限制性片段长度多态性分析)、PCR-SSO(PCR产物的序列特异寡核苷酸探针杂交)、PCR-SSP(序列特异性引物的PCR扩增)和PCR-SBT(序列直接分型)等。进行等位基因分型后,发现原先属于同一个血清学特异性的抗原往往可被数个甚至数十个不同的等位基因所编码,表明同一个特异性的HLA抗原实际上由多个亚型组成。如编码HLA-A2抗原的等位基因数已经超过100个;编码HLA-DR4的等位基因也至少有36个。
为此,HLA命名委员会制定了如下的HLA命名原则
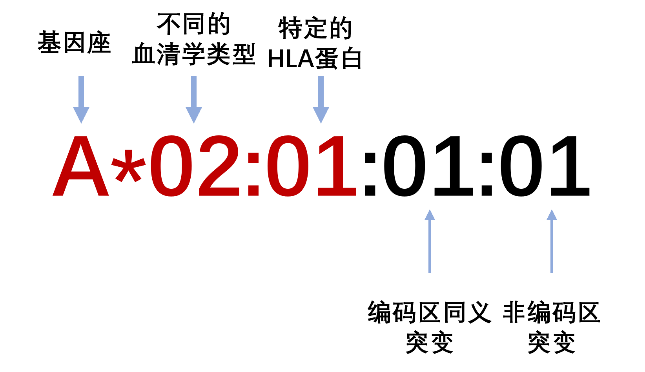
一个完整的HLA等位基因由8个数字和1个字母组成,如图:HLA-A * 02:01:01:01,其中“A”代表基因座位,“02”代表对应的血清学特异性,“01”代表一种特定类型的蛋白,“01”代表编码区碱基突变而编码氨基酸不变,第二个“02”代表内含子或非编码区碱基突变。最后两组数字是在基因上的一些微小变化,对表达的蛋白没有影响,因此可以忽略。每个人有两个HLA基因,一条来自父亲,一条来自母亲。大部分情况这两个基因是不同的(简称:杂合子);少数人有两条相同的HLA-A基因(简称:纯合子)。
举两个例子:
HLA分型1:HLA-A*03:01:01:01 HLA-A*04:06:01:01
这是杂合子,有两条不同的HLA-A基因:第一条是03型,01亚型(简称0301);第二条是04型,06亚型(简称0406)
HLA分型2:HLA-A*11:01:01:01 HLA-A*11:01:01:01
这是纯合子,两条HLA基因相同,都是11型,01亚型(简称1101)
HLA的纯合子和杂合子
纯合子:指同源染色体在同一基因座上的两个等位基因相同的基因型个体。一对相对基因有AA和aa两种基因型的个体
杂合子:指二倍体中同源染色体同一位点上的两个等位基因不相同的基因型个体,如Aa。杂合基因型比纯合显性或纯合隐性基因型的适应性都要高,这种现象被称为杂合子优势。
当位于一对同源染色体上的相同基因座位的两个等位基因中的一个(或其中部分核苷酸片段)发生缺失,与之配对的染色体上仍然存在。根据这个基因的特点,杂合性缺失表现出来的性状也会不同,如果这个缺失基因是隐性基因,杂合子或杂合性缺失后性状不会发生改变,因为未缺失的另一个等位基因不论是显性还是隐性都不会因为缺失的隐性等位基因而最终使性状改变,如果这个基因是显性基因,那么这种缺失可能表现出来,也可能不表现出来,这取决于另一等位基因是显性还是隐性。


